Reports: ND253241-ND2: Development of U Isotopes as a Tool for Reconstructing the Extent of Global Seawater Anoxia
Katharine Maher, PhD, Stanford University
The redox state
of the oceans is intricately linked to the global carbon cycle. Knowledge of
the history of marine redox conditions informs our understanding of
evolutionary pattern in the fossil record and may provide critical boundary
conditions for reconstructing depositional environments.Because
the timing and extent of past anoxia in the global oceans are poorly
quantified, the links between anoxia, impoverished marine ecosystems, and
perturbations to the global carbon cycle remain speculative.This
project is focused on (1) evaluating the use of U concentrations, [U] and d238U measured in carbonate rocks as indicators of ocean anoxia,
and (2) development of d238U
records that span Latest Permian to Late Triassic time in order to determine how
global paleoredox conditions changed following the
end-Permian mass extinction, the most
severe biotic crisis in the history of animal life.
Uranium in marine carbonate
rocks provides a unique measure of past oxygenation because [U] and d238U
in seawater depend on redox conditions. Uranium occurs in two redox states in
natural waters: U(VI) and U(IV). Under oxygenated
conditions U(VI) is abundant as the uranyl ion (UO22+). In the absence of
oxygen, U(VI) is reduced to U(IV), which is poorly
soluble and precipitates as uraninite in sediments
(UO2) (e.g. Langmuir
(1978)). Moreover, 238U is preferentially reduced from U(VI) to U(IV) (Bigeleisen, 1996),
leaving the residual U(VI) in seawater with a lower d238U value
relative to precipitated uranium in sediments (Brennecka
et al., 2010). Consequently, a shift toward more reducing conditions in the
oceans should drive simultaneous decreases in [U] and d238U.
To date we have measured [U]
and d238U in Upper
Permian through Upper Triassic limestone samples from three stratigraphic
sections arrayed along a depth transect at the Great Bank of Guizhou (GBG), an isolated carbonate platform in the Nanpanjiang Basin of south China (eastern Tethys). To further
determine whether variations in [U] and d238U on the
GBG reflect local versus global controls, we analyzed samples from the AladagNappe, Turkey (western
Tethys). Our results show a large
and abrupt decrease in d238U and [U]
at the PTB from relatively high Latest Permian values to sustained low values
in the Early Triassic (Fig. 1), and stable positive values in the Middle
Triassic (Fig. 2).
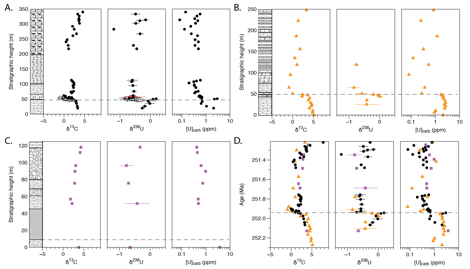
Figure 1. Stratigraphy, d13C, d238U, and [U]
for the Late Permian to Early Triassic (Induan)
interval at A) Dajiang (GBG; black filled circles), Dawen (GBG; Brennecka et al.,
2011 (unfilled circles) and this study (red filled circles); B) Taskent; C) Guandao (GBG); and D)
all locations. Gray dashed line indicates PTB. Error bars are 2 SD on replicate
analyses.
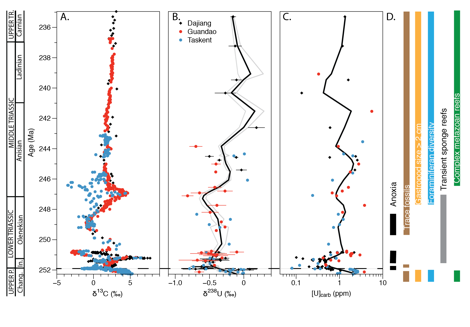
Figure 2. A)
d13C,
B) d238U
and C) [U] from the Late Permian to Late Triassic time interval. Error
bars are 2 SD of replicate analyses. Gray box shows time interval of data in
Figure 1. D) General trends in fossil records (Chen and Benton, 2012). The black fit lines are made using a
Monte Carlo-resampling scheme and the spline.smooth
function in R. Error envelopes are 2
SD.
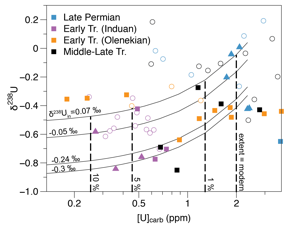
Coupled d238U and U
records allow us to constrain the uranium cycle in the Latest Permian-Late
Triassic oceans using an isotope box model. Specifically, we can directly link
changes in d238U and [U]
to the area of seafloor impacted by anoxia. We estimate that expansion of
low-oxygen conditions during the early Early Triassic
(Induan) constitutes a 25-fold increase (Fig. 3). In
addition, the period of low-oxygen conditions coincides with the large carbon
isotope perturbations (Fig. 2). The results and conclusions from this study
allow us to hypothesize that the delay of a full biotic recovery for 5 Myr until the Middle Triassic can be partially explained by
globally extensive bottom water anoxic conditions. In addition to suggesting a
link between carbon perturbations, anoxia, and suppressed biodiversity, this work
has important implications for the application and interpretation of d238U
measurements in carbonates. The reproducibility of d238U across
depositional environments and at multiple locations lends support to the global
nature of this proxy to record seawater oxygen levels given careful
consideration of diagenetic alteration.
Figure 3. d238U and
[U] with contours of different input d238U compositions.
Dashed lines indicate model predications for extent of seafloor anoxia. Open circles = Dajiang, squares = Guandao,
triangles = Taskent.
Our efforts in the coming
year will focus on (1) preparing publications, and (2) continuing to develop a
reactive transport model for early diagenesis that
enables us to link the microbial processes to the uranium and carbon isotopic
composition of key phases. The
later work will further our understanding of the U isotope dynamics at the
sediment-water interface and how their linkages to the marine carbon cycle.
References:
Bigeleisen, J., 1996. Nuclear Size and
Shape Effects in Chemical Reactions.
Isotope Chemistry of the Heavy Elements. J. Am. Chem. Soc 118,
3676-3680.
Brennecka, G. A., A. D. Herrmann, T. J.
Algeo, and A. D. Anbar (2011a), Rapid expansion of oceanic anoxia immediately
before the end-Permian mass extinction, Proceedings of the National Academy of
Sciences, 108(43), 17631-17634.
Chen, Z.-Q., Benton, M.J., 2012. The timing
and pattern of biotic recovery following the end-Permian mass extinction.
Nature Geoscience 5, 375-383.
Langmuir, D., 1978. Uranium
Solution-Mineral Equilibria at Low Temperatures with Applications to
Sedimentary Ore Deposits. Geochimica et Cosmochimica Acta 42, 547-569.
Payne, J.L., Claptham, M.E., 2012.
End-Permian mass extinction in the oceans: An ancient analog for the 21st
century? Annual Review of Earth and Planetary Sciences 41.
 printer friendly
printer friendly