Reports: ND252142-ND2: A Potential Fungal Contribution to the Selective Preservation of Long-chain Hydrocarbon Functionality in Soils and Sediments
Neal E. Blair, Ph.D., Northwestern University
The preservation of
long-chain aliphatic functional groups in sediments and soils is a common
observation that is especially pertinent to petroleum generation. The
mechanisms by which the preservation occurs are not resolved in all cases
however. This project tests that hypothesis that bound lipids in fungal
necromass may survive the initial phase of rapid diagenesis to potentially
contribute to sequestered alkyl groups.
Necromass of Fusarium avenaceum, a
common temperate soil saprotroph, was placed in stainless steel (100μm
mesh) packets and incubated in laboratory macrocosms and in the field. The
field site was the Dixon Prairie of the Chicago Botanic Garden. The contents
were analyzed via FTIR, thermochemolysis GC-MS using tetramethyl ammonium
hydroxide (TMAH), and elemental analysis-isotope ratio mass spectrometry. Fungal
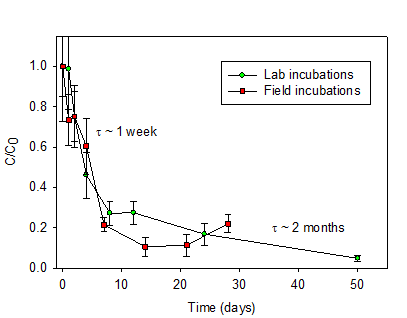
material was rapidly lost as indicated by bulk C mass (Fig. 1). Approximately
75-80% of the Fusarium OM has a
turnover time of ~ 1 week whereas ~10% has a potential turnover time exceeding
several months.
Fig. 1: The loss of bulk Fusarium OC as a function of time in soil degradation experiments
(lab and field incubations). Values are normalized to the time 0 benchmark.
Turnover times (τ) are noted for different parts of the degradation
curves.
The
selective removal of reactive fractions over time transforms the bulk and
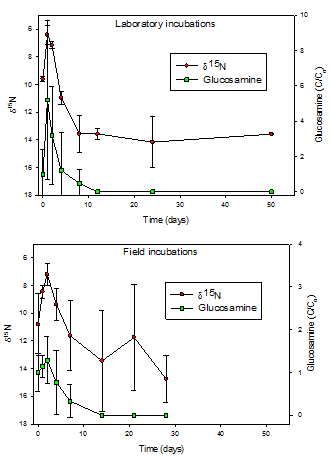
molecular composition of the fungal materials. As an example during the first
few days of incubation, the increase in concentration of glucosamine, the
monomer building block of chitin and chitosan, reflects the selective removal
of more reactive compounds (Fig. 2). Loss of the chitin/chitosan is apparent
thereafter. The parallel behavior of the d15Ntracks
the sequential removal of reactive N-containing compounds and chitin/chitosan
and suggests that the N-isotopic composition of the chitin/chitosan is
distinctly different (15N-depleted) from other nitrogenous
components.
Fig. 2: d15N and glucosamine
concentrations (C/C0) in Fusarium
necromass as a function of time of degradation in both laboratory and field
experiments (n=3). Note that the d15N scale is inverted to
better illustrate the parallel behaviors.
FTIR analysis also
reveals changes in organic composition over time. The reduction in the
absorbance of a C-O stretch centered at 1073 cm-1 is likely due to
the loss of a dominant carbohydrate, such a β-glucan or chitin/chitosan.
Additional C-O containing components as well as amide functionality persist to
50 days, though with changes in their relative abundance. Insofar as chitin
appears to be effectively removed during degradation (Fig. 2), we conjecture
that the residual material may be a cross-linked cell wall glycoprotein or its
diagenetically altered product. Changes in the relative concentrations of
methylated hexoses derived from TMAH-thermochemolysis also indicate
preferential loss of some carbohydrates.
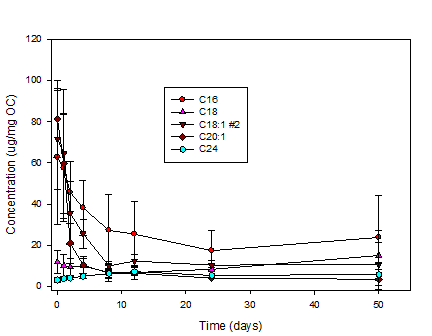
Long-chained n-fatty
acids exhibit a range of reactivities despite their similar structures (Fig. 3).
The order of reactivity is C20:1> C18:1>C16> >C18>C24. Notable
quantities of the fatty acids persist to the end of the experiment. We
hypothesize that the fatty acids reside in and/or are bound to biochemical
matrices with varying turnover times.
Fig. 3: Fatty acid
concentrations in degrading Fusarium as
a function of time in laboratory degradation experiments.
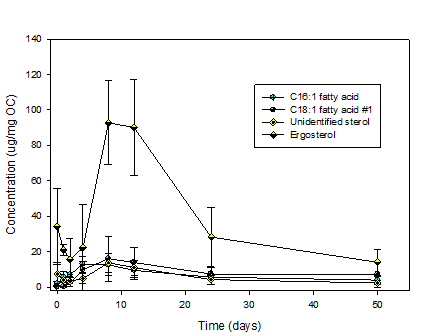
Fig. 4: Fatty acid
and sterol concentrations in degrading Fusarium
as a function of time from the laboratory experiments.
Other lipids exhibit
a more complex behavior (Fig. 4). The fungal biomarker ergosterol and an
unidentified sterol decrease rapidly at first and then peak at days 8-12. The
n-fatty acids C16:1 and C18:1 (a different isomer than that depicted in Fig. 4)
reach maximum concentrations at days 4 and 8 respectively and then decrease.
Fungi contain sterol-rich domains, such as chitisomes, that are perhaps
concentrated in the early stages of degradation and then decomposed.
Alternatively, colonization of the Fusarium
OM via other soil fungi may be contributing to the lipids. Microscopic and SEM analyses
of the residue do indicate hyphae and spores from other fungi however the bulk
of the material appears to be the Fusarium
residue.
The tentative
conclusion reached from these data is that some lipid fractions may persistent
long enough to contribute long-term sequestration, especially if cross-linking
or other diagenetic reactions render them biologically unreactive. We are
currently analyzing samples that used other fungal species and are yielding
fundamentally the same results. The identity of the persistent macromolecular
material that is serving as a host for the lipids is also being sought. The
role of fungi in the sequestration of C in soils and sediments has barely been
considered, thus this study should open new avenues of research in the field.
This project has
allowed me to move into several new areas of research involving microbial
cycling in soils, the contribution of fungi to the C-cycle and the use of
biomarkers in organic geochemistry. This has led to proposal submissions to NSF
and DOE. This was a new direction as well for the post-doctoral fellow
supported by the project, Kathryn Schreiner, who plans to continue this work as
an assistant professor at the University of Minnesota – Duluth. This project
has also provided research opportunities for undergraduate students. Jessie
Moravek is studying the potential role of chitin as a source of bound fatty
acids. Aarohi Shah is studying lipid production by chitinolytic
fungi. Both students have had to do extensive method development and have been
remarkably successful. We have also hosted a REU student from the Chicago
Botanic Garden (Ben Sedillo) this summer who began studies with additional
fungal species. He was introduced to fungal biochemistry and FTIR analyses for
the first time.
 printer friendly
printer friendly