
ACS PRF | ACS
All e-Annual Reports

41183-GB2
Dominican Republic Corals as High-Resolution Records of Shallow Marine Paleoenvironmental Conditions Prior to Closure of the Panamanian Seaway
U-Pb DATING A MIOCENE CORAL
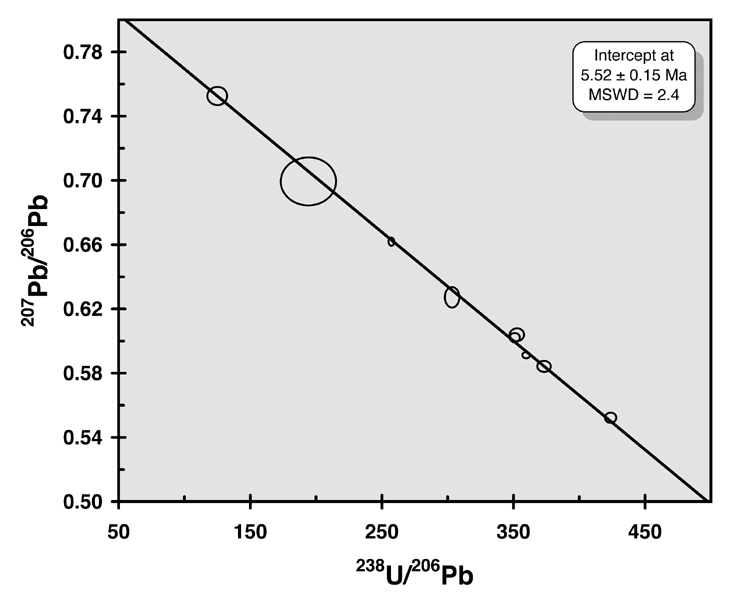
Fig. 1. 207Pb/206Pb - 238U/206Pb 3D inverse linear concordia diagram for a Miocene Caribbean coral.
Closure of the Central American Seaway (CAS) in the Pliocene isolated the Caribbean and Pacific basins and contributed to a pulse of extinction and origination of species of shallow marine fauna on both sides of Central America (Jackson et al., 1996). Studies aimed at calculating evolutionary rates associated with this event require not only detailed paleontological assessment, but also robust chronologies of sedimentary units deposited prior to, during, and after CAS closure. One limitation to such studies has been insufficient age control on some fossil-bearing lithologies that has introduced significant uncertainty into the rates of change of marine communities (McNeill et al., 2000). The Cibao Valley of the Dominican Republic contains an ~3 My-long record of well-preserved fossil taxa that existed in the Caribbean prior to final closure of the CAS, and one well studied unit in this area is the Gurabo Formation, which was deposited prior to final closure of the CAS. However, the precise age of the Gurabo Formation remains the subject of debate; in fact, over the past 90 years, 19 studies have reported various and disparate ages for Cibao Valley units, with the ages of the Gurabo Formation ranging from the middle Miocene (~10 Ma) (Ramirez, 1956) to the middle Pliocene (~3 Ma) (van den Bold, 1972).
Shallow marine sequences can be difficult to categorize chronostratigraphically given highly variable deposition rates, reworking of sediments, lack of age diagnostic fossils, and diagenetic alteration (Getty et al., 2001). Portions of the Gurabo Formation are composed of low permeability muds, and as a result, some corals remain largely pristine. We identified an extremely well preserved coral head (of the now extinct species Goniopora hilli) and dated it using 238U/206Pb -- 207Pb/206Pb techniques.
Coral subsamples were carefully screened for signs of diagenetic alteration. Nine splits of the samples that passed these screening tests were dissolved, spiked with a mixed 236U-233U-205Pb tracer, processed using standard column chemistry techniques (Manhes et al., 1978), and analyzed by TIMS and MC-ICP-MS. A 238U/206Pb -- 207Pb/206Pb 3-D inverse linear concordia age of 5.52 ± 0.15 Ma (2s) (Figure 1) was calculated using Isoplot (Ludwig, 1996). In addition, Sr isotopic ratios on four subsamples yield an average 87Sr/86Sr ratio of 0.708993 ± 0.000029 (2s) which intersects the global marine Sr curve within error of the U-Pb age. This chronology opens the door for more direct absolute age determinations in this sequence and thus tighter constraints on rates of species origination prior to closure of the CAS.
SEA SURFACE TEMPERATURE AND SALINITY VARIABILITY
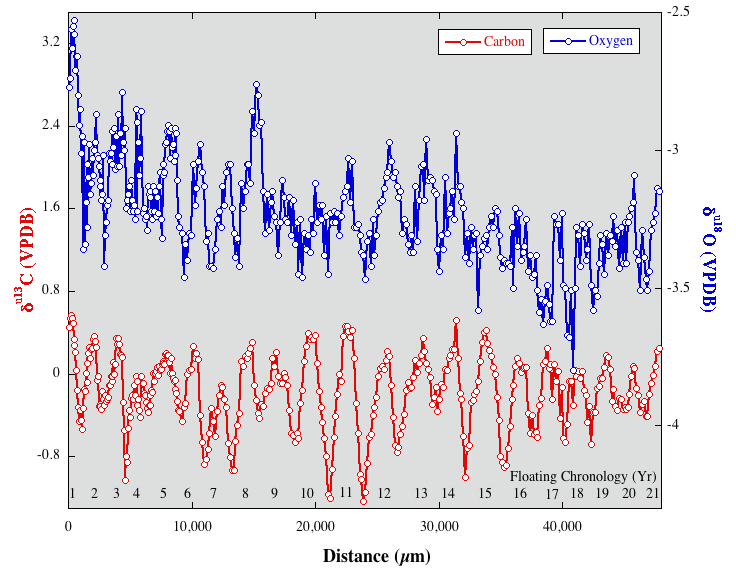
Fig. 2. Oxygen (blue) and carbon (red) isotopic profiles from G. hilli showing well-defined seasonality.
Aside from age uncertainties in important sedimentary sequences, we lack a clear understanding of how shallow Caribbean environments changed in response to closure of the CAS. This is particularly true for changes at the seasonal scale, a time frame that plays an important role in determining the composition of shallow marine communities (Ateweberhan et al., 2006). A 21-year-long, high-resolution stable isotope and trace element record of a late Miocene (~5 Ma) coral from the Dominican Republic was constructed that records shallow marine paleoenvironmental conditions prior to final closure of the CAS. This coral serves as one snapshot for the late Miocene central Caribbean that, when integrated with other records, may allow a better understanding of the role played by environmental variables in forcing Neogene faunal turnover in the Caribbean.
A single corallite from the same coral head of G. hilli was isolated and micromilled at 150 µm intervals for stable isotopic analysis. When plotted versus distance along the corallite growth axis, carbon and oxygen isotopic ratios define clear and quasi-regular sinusoids (Figure 2). Oxygen isotopic ratios are controlled primarily by water temperature and oxygen isotopic composition. Seasonal ranges in d18O values of the Gurabo G. hilli average 0.4ä ± 0.1ä (1s) (Figure 2), a value that if attributed solely to water temperature, corresponds to an average seasonal range of ~2űC, nearly identical to modern values from Haiti (Slutz et al., 1989). However, seasonal or long-term changes in ocean water d18O values and salinity can mask temperature signals in coral d18O values (Swart et al., 2001). Combining Sr/Ca and oxygen isotopic ratios can allow parsing salinity and water temperature variations (Ren et al. 2002). However, Sr/Ca analyses were performed on splits of powders used for stable isotopic analysis, but the results suggest that applying Recent Goniopora Sr/Ca -- temperature calibrations to a Miocene G. hilli introduces such a high degree of uncertainty that the Sr/Ca-based temperature reconstructions are of little use.